|
||
Класс Трематоды, или Сосальщики (Trematoda = Digenea)Царство Животные (Zoa), Подцарство Многоклеточные (Metazoa), Надраздел Эуметазои (Eumetazoa), Раздел Двусторонне-симметричные (Bilateria), Тип Плоские черви (Plathelminthes), Класс Трематоды, или Сосальщики (Trematoda = Digenea) Трематоды — паразитический класс плоских червей. Сохраняя все основные признаки типа, они характеризуются специфическими особенностями, которые обусловлены их эндопаразитическим образом жизни. Прежде всего это наличие специальных органов прикрепления — присосок и замена ресничного эпителия лишённым ресничек тегументом. Пищеварительная система трематод, как правило, развита хорошо, но имеется тенденция к её рудиментации и исчезновению. Одновременно с этим приобретает особо важное значение способность трематод к восприятию пищевых веществ извне, через покровы. Как и другие паразитические организмы, трематоды разных поколений производят огромное количество яиц и личинок. Трематодам присуща также тенденция к живорождению, полностью осуществлённая на некоторых этапах их жизненного цикла. Жизненный цикл трематодСам по себе жизненный цикл трематод, связанный с чередованием поколений, совершенно не свойствен свободноживущим плоским червям и возник в ходе эволюции, как следствие перехода этой группы к паразитизму.
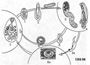
Жизненный цикл трематод представляет собой одно из самых удивительных явлений живой природы. Он очень сложен, проходит с участием нескольких хозяев, сопровождается чередованием поколений и сменой паразитических и свободноживущих фаз развития. Половозрелая гермафродитная особь — марита паразитирует во внутренних органах позвоночных животных. Откладываемые ею яйца выводятся из организма хозяина во внешнюю среду. Это сложные яйца, под плотной скорлупкой которых находится не только яйцеклетка, но и множество желточных клеток, поставляющих необходимые развивающемуся зародышу питательные вещества. В типичном случае яйцо для завершения своего развития должно попасть в воду, и там из него выходит подвижная личинка — мирацидий. За время короткой жизни эта личинка должна проникнуть в первого промежуточного хозяина — моллюска определённого вида, в котором претерпевает превращение (метаморфоз), связанное с упрощением организации. В результате развивается половозрелая фаза — материнская спороциста. Это очень просто устроенный организм, чаще всего мешковидной формы, способный к размножению путём партеногенеза. Партеногенетические яйца материнской спороцисты, обычно называемые генеративными или зародышевыми клетками, дробятся без оплодотворения и дают начало новому, дочернему поколению — редиям. Последние, покидая материнский организм, активно ползают между органами моллюска хозяина. Обычно они обосновываются в его кровеносных лакунах (кровеносная система моллюсков не замкнута) или в пищеварительной железе. Редии, как и материнские спороцисты, размножаются только партеногенетически, отрождая либо новые поколения редий, либо своеобразных хвостатых церкарий, которые представляют собой личинок гермафродитного поколения трематод (мариты). Редии устроены сложнее, чем материнские спороцисты, так как имеют хорошо развитую пищеварительную систему, представленную мускулистой глоткой и кишечником. Второе партеногенетическое поколение — обязательное звено в жизненном цикле трематод, но у многих высших трематод в материнских спороцистах развиваются не редии, а морфологически иные, более просто устроенные партеногенетические организмы — дочерние спороцисты. Об их соотношении с редиями будет рассказано ниже. Здесь ограничимся лишь сведениями о том, что дочерние спороцисты лишены пищеварительной системы и имеют обычно мешковидную или шарообразную форму тела. Они редко отрождают себе подобных, как правило, давая начало лишь церкариям. Все партениты — партеногенетическое поколение трематод — живородящи. Церкарии представляют собой очень подвижных маленьких личинок, снабжённых специальным органом движения — хвостом. Выйдя из тела родительской особи, они покидают организм своего хозяина-моллюска и попадают в воду. Здесь судьба их может быть различной. Церкарии одних видов довольно быстро оседают на прибрежной траве или на заглатываемых нередко птицами подводных предметах (песчинки, мелкие камешки, пустые раковинки моллюсков и тому пподобном). Они одеваются плотной защитной оболочкой (инцистируются) и превращаются в покоящуюся фазу, называемую адолескарией. Будучи случайно проглочена каким-либо позвоночным животным — окончательным хозяином данного вида трематод, личинка выходит из цисты (эксцистируется) и превращается в мариту. Церкарии других видов для дальнейшего развития должны попасть в организм второго промежуточного хозяина (иногда его называют дополнительным). Чаще всего это головастики, мелкие рыбки, личинки водных насекомых, моллюски, пиявки и другие. В организме второго промежуточного хозяина церкарии обычно инцистируются, переходя в фазу метацеркарий. Если зараженный второй промежуточный хозяин будет съеден позвоночным животным, подходящим для развития метацеркарий данного вида, то личинка выходит из цисты и достигает в окончательном хозяине половой зрелости, превращаясь в гермафродитную особь. Таким образом, в жизненном цикле трематод обязательно участвуют три поколения — два партеногенетических и одно гермафродитное. При этом происходит смена двух разных типов полового размножения: обоеполого, связанного с обязательным оплодотворением яйцеклетки, и партеногенетического, которое осуществляется без оплодотворения. В отличие от метагенеза, при котором половое размножение чередуется с бесполым, чередование поколений, свойственное трематодам, называется гетерогонией. Онтогенез, то есть развитие организма от стадии яйца до половозрелости, у всех трёх поколений трематод проходит различно. Онтогенез первого партеногенетического поколения — материнской спороцисты связан с метаморфозом. Её личинкой является мирацидий, который развивается из оплодотворённого яйца мариты. Второе партеногенетическое поколение — будь то редия или дочерняя спороциста — развивается без метаморфоза. Третье, гермафродитное, поколение опять развивается с метаморфозом, но имеет не одну, а две личинки: церкарию и метацеркарию, или адолескарию. Характерно, что первым промежуточным хозяином трематод всегда служит моллюск. Роль второго промежуточного хозяина могут выполнять самые разные животные — от кишечнополостных и гребневиков до млекопитающих включительно. Окончательный же хозяин — позвоночные. По отношению к первому промежуточному хозяину трематоды проявляют узкую специфичность. Это означает, что материнские спороцисты каждого конкретного вида трематод могут паразитировать в моллюсках лишь одного или нескольких близких видов. Напротив, ко второму промежуточному хозяину трематоды, как правило, очень широко специфичны. Метацеркарии одного вида могут поселяться в животных, относящихся не только к разным видам, но и к разным классам и даже типам. Специфичность, вернее приуроченность к тому или иному виду животных — хозяев, объясняется здесь экологическими причинами. Дело в том, что роль второго промежуточного хозяина конкретного вида трематод выполняют лишь те животные, которые служат пищей окончательному хозяину данного вида. Это важное условие для осуществления жизненного цикла паразита. По образному выражению известного немецкого учёного А. Лооса, окончательный хозяин, выискивая свою «излюбленную» пищу, как бы сам подыскивает и собирает своих паразитов. Разумеется, из этого правила могут быть исключения: известны трематоды, которые и на стадии метацеркарий могут быть узкоспецифичны к своему хозяину. Следует ещё сказать несколько слов о взаимоотношениях между трематодами и моллюсками. Выше говорилось об узкой специфичности трематод к их первому промежуточному хозяину. Обращает на себя внимание и удивительная сбалансированность отпошений между партнерами системы паразит — хозяин. Зараженный моллюск, несмотря на наличие паразитов, может долго сохранять жизнеспособность. Морфология и физиология маритыОбычно когда говорят о трематодах, имеют в виду паразитирующую в позвоночных животных мариту половозрелую фазу гермафродитного поколения. Ограничиться описанием только одной мариты, как это часто делается, было бы неверно. Мы уже знаем, что трематоды характеризуются сложным жизненным циклом, который включает по крайней мере три резко отличающихся друг от друга поколения. Поэтому представить особенности строения и биологии трематод можно, только рассматривая каждое из поколений в отдельности. Наиболее сложно устроена марита, которая изменена паразитическим образом жизни менее других поколений и во многом сходна с организацией остальных плоских червей. С описания её строения и биологии мы начнём наше знакомство с трематодами.
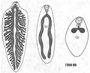
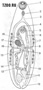
Тело мариты по большей части имеет уплощённую листовидную форму и несёт одну или две присоски Покровы трематод представлены типичным тегументом. Его поверхность, лишённая ресничек, часто несёт переплетающиеся гребни и складки, которые играют существенную роль в питании (смотри ниже). Слои кольцевых и продольных мышц хорошо развиты. Паренхима имеет типичное для плоских червей строение, но, в отличие от паренхимы свободноживущих турбеллярий, выполняет ещё и роль депо гликогена, который служит для трематод основным источником энергии (смотори ниже).
Пищеварительная система начинается ротовым отверстием, которое ведёт в округлую мускулистую глотку, играющую роль насоса при заглатывании пищи. За глоткой следует пищевод. Он разделяется на две кишечные ветви, заканчивающиеся слепо. У крупных трематод, например у печёночной двуустки, кишечник обильно разветвлён.
Половая система трематод устроена сложно и, как у всех плоских червей, гермафродитна.
Матка служит для выведения яиц наружу. Но у многих видов (отряды Plagiorchiida, Cyclocoelida и другие) в яйцах, ещё находящихся в матке, начинается дробление овоцита, а нередко проходит и всё эмбриональное развитие личинки, так что наружу выходят яйца, в которых заключён вполне готовый мирацидий. Это очень выгодно в биологическом отношении, так как развивающиеся эмбрионы полностью избавлены от неблагоприятных воздействий внешней среды (резкие колебания температуры, высыхание и так далее). В результате процент выживающих яиц паразита значительно возрастает. Однако эмбриогенез требует определённого времени, и развивающиеся яйца задерживаются в петлях матки, а так как поступление новых яиц непрерывно продолжается, матка становится очень длинной и её многочисленные петли занимают иногда до 2/3 тела червя. Несмотря на то, что половая система мариты гермафродитна, самооплодотворение встречается очень редко, что невыгодно для вида, так как даёт генетически однородное потомство. Поэтому оплодотворение, как правило, перекрёстное. Известны даже случаи появления вторичной раздельнополости марит. Таковы, например, многие представители своеобразных паразитов морских рыб из семейства Didymozoidae. Полностью раздельнополым становится всё семейство Schistosomatidae — паразитов кровеносной системы человека и животных. Более крупный самец носит самку в особом «канале», который образован свёрнутыми в широкую трубку краями его листовидного тела. Мариты трематод могут паразитировать в любых тканях и органах позвоночных животных. В зависимости от условий существования в тех или иных органах хозяина возникают соответствующие изменения строения и физиологии паразита. Так, мариты, поселяющиеся в кровеносных сосудах или в каких-либо узких протоках (например, желчных протоках печени), отличаются длинным, нитевидно вытянутым телом. В то же время многие тканевые паразиты имеют почти шаровидную форму. Тесно связано с местом поселения и строение органов прикрепления мариты. У паразитов кишечника, особенно у видов, обитающих в прямой кишке, откуда они легко могут быть выброшены при физиологических отправлениях хозяина, присоски развиты особенно сильно. Напротив, у трематод, паразитирующих в полости тела (например, у паразитов из семейства Cyclocoelidae), присоски дегенерируют. От локализации нередко зависит и способ питания мариты. Так, тенденция к замене активного питания всасыванием растворённых органических веществ через покровы особенно заметно проявляется у паразитов крови. Мариты позвоночных нередко оказываются в условиях острого недостатка кислорода. Это влияет на характер обмена, который приближается к анаэробному, так как нормальное дыхание в большинстве случаев оказывается невозможным. В разных органах позвоночного животного парциальное давление кислорода может быть совершенно различным. С этим связано то обстоятельство, что трематоды не утратили способности использовать кислород, если он имеется. Поэтому им свойствен не чисто анаэробный (как предполагали раньше), а смешанный тип обмена. В зависимости от того, в каких органах поселяются мариты, в их обмене могут преобладать либо анаэробные, либо аэробные процессы. Те виды, которые паразитируют в трахее или в лёгких хозяина, способны к нормальному дыханию. Мариты, питающиеся кровью, по-видимому, могут использовать кислород, связанный гемоглобином крови хозяина. Наконец, в тканях многих трематод содержится гемоглобин, отличающийся повышенным сродством к кислороду. Это позволяет маритам использовать даже то незначительное количество свободного кислорода, которое имеется в среде их обитания. Трематоды, как и другие эндопаразитические черви, получают необходимую для жизнедеятельности энергию в результате расщепления гликогена, содержащегося в их тканях. Жиры не могут служить для них источником энергии, так как их расщепление возможно только в присутствии свободного кислорода. В тканях паразитических червей заключены огромные запасы гликогена. У свободноживущих родичей трематод таких запасов гликогена никогда не бывает. Прежде всего это обусловлено тем, что свободноживущие организмы широко используют в качестве источника энергии жиры. С другой стороны, анаэробное расщепление гликогена — процесс крайне невыгодный в энергетическом отношении. Если в присутствии кислорода гликоген расщепляется до углекислого газа и воды, то в анаэробных условиях он распадается на крупномолекулярные соединения (главный продукт обмена — молочная кислота), при этом высвобождается в 10 раз меньше энергии, чем в первом случае. Вот почему так велики в теле мариты запасы гликогена, составляющие до 60% её сухой массы. Не следует забывать, что много гликогена расходуется на образование бесчисленных яиц этих червей.
Т. А. Гинецинская. Класс Трематоды, или Сосальщики. Жизнь животных. Том первый / Гл. ред. В. Е. Соколов -Москва: Просвещение, 1987 |
||
|
© Царство животных. Since 2006 |
||